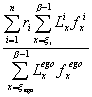Non penso si possa venire tacciati
di sciovinismo settoriale se si afferma che l'oggetto della demografia è uno
dei più interdisciplinari tra quelli delle scienze sociali. Questo però non
è merito (se vogliamo considerarlo tale) dei demografi, bensì dell'argomento
"popolazione" in sé, che si situa appunto al crocevia tra diversi ambiti[2]:
vi si intersecano biologia, antropologia, storia, sociologia ed economia (per
tacere della statistica, della quale la demografia nell'opinione di molti
non rappresenterebbe molto più che una branca). Non sembra, infatti, che queste
contiguità raccolgano grande interesse[3] — mi riferisco in particolare a quella
con la biologia[4].
Il cuore degli studi demografici è
oggi infatti orientato verso l'aspetto statistico e modellistico ("a point of proper disciplinary pride" secondo
l'espressione di McNicoll, citato in de Bruijn, p.1)[5]
— comprensibilmente, d'altronde, visto l'interesse degli sviluppi e delle
applicazioni dell'impianto della popolazione stabile e di altri raffinati
metodi di analisi, amplificati della potenza che l'uso dei computer conferisce
loro. Troverei però illogico sprecare questo potenziale di eclettismo[6],
pur con tutti i rischi del caso[7]: sono proprio
le conquiste delle singole discipline, che rendono più attraente la specializzazione,
a dare urgenza anche a uno sforzo di raccordo. Senza una riflessione generale,
rischiamo di rompere la conoscenza scientifica in un insieme di cocci[8], per quanto luccicanti — e questo può portare
guai ben più a lungo che per sette anni.
Cominceremo dunque col passare
in rassegna alcuni punti ove la demografia più aiuta a comprendere il nesso
tra esistenza umana e sostrato biologico[9]. Una discussione
così generale su metodi ed interpretazioni darà poi lo spunto per alcune eterogenee
considerazioni di ordine filosofico, presentate nella seconda parte.
1 Biologia
Procedendo dal particolare al
generale, il primo compito della demografia — coerente d'altronde con l'impostazione
statistica che essa è venuta ad assumere — è quello della raccolta dei dati.
Il monitoraggio delle tendenze
presenti si accompagna poi alla ricostruzione di quelle passate ed alla previsione
di quelle future. Questa funzione ancillare può rivelarsi preziosa per altre
scienze sociali, sempre si riesca a comunicare attraverso le frontiere che
dividono i campi del sapere, e può implicare una grande raffinatezza tecnica.
Vi è infine il momento interpretativo,
che non può essere appannaggio esclusivo della demografia, così come di qualsiasi
altra impostazione (È una questione di buon senso che una molteplicità di
punti di vista possa dare frutti più copiosi).
In ambedue i pilastri della disciplina,
l'analisi della mortalità e quella della fecondità (a sua volta collegata
a fertilità[10] e nuzialità),
vi è un evidente collegamento con i due motori principali della selezione:
la sopravvivenza e la riproduzione. Le esigenze di esposizione inducono ad
esaminare un punto alla volta, ma non va dimenticato che uno dei contributi
fondamentali della demografia è appunto lo studio delle interrelazioni tra
fenomeni diversi.
1.1 Il metodo
Vi è un ovvio uso della statistica
nella genetica di popolazione, che può anzi considerarsi un'applicazione particolare
del calcolo delle probabilità, basato sulle frequenze geniche e sulle leggi
biologiche. E da questo punto di vista vi sarebbe già una certa parentela
di metodo con una parte importante dell'analisi demografica, pur con importanti
differenze[11].
Possiamo anzi affermare con Scardovi
(1994, p.202): "Demografia e biologia
trovano la più immediata intersezione nella parte in cui la genetica di popolazioni
si rivolge alle popolazioni umane: l'una è attenta agli individui, l'altra
ai geni di cui sono portatori. L'intreccio fenomenologico è dunque in re
ipsa: non c'È evento osservato in demografia
che non abbia un'origine o una ripercussione biologica"[12]
(si vedano anche Lucchetti e Soliani, p.159, e Cliquet pp.183 e 207-8). Barrai
(p.41) aggiunge che "esiste un'ampia
sovrapposizione di metodi e di oggetti di studio tra le due discipline [demografia
e genetica], che è assai vasta nel caso
della genetica di popolazioni umane. In pratica, alcuni dei problemi più importanti
della genetica di popolazioni umane possono essere risolti solo con metodi
demografici, e con l'aiuto della demografia. In particolare, la sovrapposizione
delle generazioni, la tavola attuariale, lo studio della mortalità e della
fertilità, la crescita delle popolazioni, la migrazione e la distribuzione
dei gruppi sono argomenti di studio comune".
Vi sono però punti di contatto
più specifici con la teoria dell'evoluzione; il concetto-base di fitness darwiniana,definito in Bodmer e Cavalli-Sforza come numero medio di figli (II,
p.53), è infatti di evidente natura demografica: una sorta di tasso netto
di riproduzione senza distinguere i figli per sesso[13] — tenendo conto che la base di riferimento
ideale non è una coorte di individui, ma un genotipo relativo ad un determinato
locus. L'idoneità è così funzione
tanto della fecondità quanto della sopravvivenza differenziali; da questo
possiamo poi derivare il coefficiente
di selezione e modelli dinamici di mutazione delle frequenze geniche nella
popolazione[14].
Concentriamoci ora su una delle
questioni centrali della sociobiologia: l'interpretazione in chiave evoluzionistica
(la logica è di selezione individuale, non di gruppo) dell'altruismo[15].
Icastica al proposito l'affermazione di Haldane, cui si chiese una volta se
avrebbe dato la vita per un fratello: "Per
un solo fratello no, ma per due sì, oppure per otto cugini"[16].
L'interpretazione sociobiologica si basa in prima istanza su un'estensione
del concetto di fitness darwiniana come era stato definito in precedenza:
"L'idoneità complessiva si misura dalla quota
di geni che un organismo trasmette alle generazioni discendenti come risultato
della sua stessa riproduzione e di quella dei collaterali consanguinei, in
quanto portatori d'una parte dei suoi medesimi geni" (Gallino p.XI, che
cita Hamilton).
Un approfondimento illustra un
possibile contributo dei metodi demografici: avrebbero i nostri generosi (quanto
avveduti) eroi sacrificato la propria vita per salvare quella di otto bisnonni?
Per rispondere dobbiamo dinamizzare l'analisi dell'altruismo anzitutto rispetto
all'età, considerando la speranza di vita residua degli individui da salvare,
e non solo il coefficiente di parentela biologica[17].
Un ulteriore raffinamento consiste nell'effettuare la ponderazione degli anni
di vita residui con la fecondità per età ed anche nel prendere in considerazione
— come usuale in demografia — la speranza di vita non fino alla morte, ma
solo fino al termine dell'età feconda[18], particolarmente
rilevante (finora) per le donne; tralasciando questioni non essenziali, quali
l'assistenza alla propria prole già in essere (che verrebbe compromessa in
caso di decesso del genitore), immaginiamo allora un rapporto benefici/costi:
ove gli n individui i per i quali
sacrificarsi condividono la porzione r
(variabile in ognuno di essi) di geni con il soggetto — si tratta dunque del
coefficiente di parentela biologica[19] — mentre
le x
indicano l'età al momento dell'osservazione[20]. Il numeratore
rappresenta il ricavo dell'operazione, il denominatore il costo: se il rapporto
sale sopra l'unità, la riproduzione del pool genico dell'individuo verrà aumentata
con il sacrificio[21].
1.2 Fecondità, fertilità e nuzialità
La semplice raccolta di dati per
la comprensione delle tendenze in atto è di estrema importanza in particolare
in questo momento: l'Italia (in misura minore anche gli altri paesi economicamente
avanzati) sta sperimentando la fecondità più bassa mai conosciuta nella storia
per un gruppo umano di proporzioni rilevanti. Anche prescindendo dall'importanza
congiunturale del fenomeno, rimane l'interesse scientifico dell'osservazione
degli effetti (e dei presupposti) di una così accentuata denatalità — occasione
particolarmente preziosa giacché la prova sperimentale non è in sostanza possibile
nelle scienze sociali.
La raccolta di dati e l'applicazione
di metodi demografici possono rivestire grande importanza anche per lo studio
della fertilità[22] (soprattutto
differenziale: ad esempio per età, ordine di nascita o durata dell'unione).
Il contributo all'analisi biologica si ritiene solitamente maggiore, quanto
più le popolazioni considerate sono lontane dal controllo delle nascite[23], come avviene negli studi antropologici
o in quelli di demografia storica.
Risulterà almeno implicitamente
dal seguito di queste pagine come la demografia possa portare un contributo
sia sul piano della validazione/falsificazione che su quello della formulazione
vera e propria di teorie. E certo l'approccio evoluzionistico può contribuire
molto a capire le dinamiche alla base di nuzialità e fecondità. Hobcraft e
Kiernan si soffermano su questo aspetto: citando anche Ridley, mettono in
rilevo come la selezione naturale non potesse distinguere tra inclinazione
alla sessualità e procreazione, inscindibili di fatto nelle specie animali.
Nell'uomo si è realizzato in tempi recenti, in particolare con la diffusione
dei metodi anticoncezionali, uno iato tra questi due aspetti[24]:
da qui sono ipotizzabili tensioni verso le quali non siamo geneticamente attrezzati[25]. Leridon, d'altra parte, mette in rilievo
la valenza selettiva del matrimonio: non si tratta solo del vantaggio dei
più sani e/o attraenti in regime di libera scelta del coniuge, ma anche di
altri meccanismi[26]; su questo si vedano anche Lucchetti e
Soliani §3.1 e Cliquet pp.187-9 (vedi anche quanto riportato in n.46).
La sezione 1.6 è dedicata alla questione della "fine della selezione naturale",
collegata anche a questi argomenti.
Una questione più specifica è
poi quella dei presupposti dell'analisi della fecondità di fronte ai cambiamenti
tecnologici in campo riproduttivo. Già i demografi hanno dovuto rivedere le
proprie assunzioni in merito alla cogenza del nesso nuzialità/fecondità (in
alcuni paesi europei metà delle nascite avviene fuori dal matrimonio): l'analisi
della nuzialità ha così perso parte della propria importanza, assurgendo però
a maggiore dignità sul piano dell'interesse autonomo. Ora poi le tecniche
di fecondazione artificiale minacciano non solo i tradizionali limiti di età
feconda femminile a e b ed il rapporto k
dei sessi alla nascita (cfr. nota 13)[27],
ma persino la coincidenza tra discendenza sociale e biologica anche per quanto
riguarda le donne. Mater semper certa,
pater numquam rischia inopinatamente l'obsolescenza per eccesso di ottimismo.
Se è quest'ultima circostanza
ad essere forse più preoccupante scientificamente (specie per quanto riguarda
il nesso demografia/evoluzionismo), va ammesso tuttavia che c'è invece da
sentirsi inquieti soprattutto di fronte alla prospettiva di una composizione
per sesso delle generazioni future in balìa delle oscillazioni della moda[28]
(anche se una volta tanto il sistema incorporebbe un premio per la categoria
meno trendy).
1.3 Mortalità e morbilità
Quest'analisi trovò già nel XVIIº
secolo uno strumento di grande potenza nelle prime tavole di mortalità di
Graunt, e costituisce oggi la colonna portante della non sempre allegra scienza
demografica.
è peraltro evidente l'interesse
che anche per le discipline biologiche può assumere un sistematico studio
statistico di mortalità e morbilità[29]. Dall'osservazione
delle cause di morte otteniamo dati preziosi per comprendere le modalità della
selezione naturale sia nel passato che in prospettiva. L'analisi differenziale
viene condotta principalmente per sesso, condizione socioprofessionale o appartenenza
nazionale[30] — oltreché
naturalmente per età, che è la distinzione fondamentale. L'estrema rilevanza
delle mediazioni sociali e culturali rende però difficile ricavarne elementi
illuminanti per comprendere il terreno sul quale la selezione naturale si
dispiega nella sua forma più eclatante, "la sopravvivenza del più adatto".
Per quanto ivi applicabili, questi metodi sono più fruibili se applicati alle
popolazioni animali.
Su questo terreno di confine tra
demografia, epidemiologia e statistica medica è possibile ad esempio valutare
la reale incidenza delle diverse cause di morte in termini di anni sottratti
alla speranza di vita (che può venire calcolata in assenza di una o più di
esse). Si vede così come non sempre quelle cui è attribuibile il maggior numero
di decessi siano quelle che di fatto sottraggono maggiore vita (misurata in
persone-anno): torneremo su questo nella sez. 2.1.
Un'altra distinzione di rilievo
è quella tra età media, età media al decesso, speranza di vita e longevità.
La prima si riferisce all'età che ad una certa data hanno i membri della collettività
in esame. La speranza di vita è il numero di anni che in media possono aspettarsi
di vivere da una certa età in poi: la più usata è la speranza di vita alla
nascita (calcolata perciò all'età 0); questa non corrisponde che in circostanze
molto particolari (stazionarietà della popolazione) all'età media al decesso,
che è influenzata dalla struttura per età. La longevità può riferirsi infine
all'età alla quale mediamente dovrebbe avvenire quella che nel linguaggio
comune si chiama morte di vecchiaia, intendendo per essa non la morte all'età più avanzata,
bensì semplicemente quella non precoce[31].
1.4 Le migrazioni
Il fenomeno è forse ancora più
importante per i biologi che per i demografi (cfr. Jacquard, p.36). Innanzitutto,
per i genetisti di popolazione — i quali però, pur avendo bisogno dei dati
dei demografi, definiscono la popolazione in modo molto diverso, in base alle
frequenze geniche. Inoltre gli spostamenti degli individui (vale anche per
gli animali) sono fondamentali per la trasmissione delle malattie infettive
— senza contare gli effetti della densità di popolazione sulla trasmissione
del contagio.
Per un'analisi generale delle
interrelazioni tra questo fenomeno demografico e la biologia (in particolare
la genetica di popolazione) rimando agli interventi di Termote, Lucchetti
e Soliani, e Cliquet.
1.5 Demografia storica
L'approccio demografico permette
di gettare luce sui comportamenti dell'uomo comune, laddove essi sono massimamente
rilevanti: nascita, matrimonio (o comunque procreazione), morte. Si realizza
così una storiografia dal basso, spesso appoggiata su una buona documentazione,
opposta a quella di impronta ottocentesca — attenta alle vicende di pochi
grandi individui (quando non anche alle inclinazioni delle "razze").
Nell'ambito della storia demografica
appare di particolare interesse la distinzione introdotta nella sezione 1.3:
quando si legge che la speranza di vita nelle società preindustriali era intorno
ai 25-30 anni[32], viene spontaneo
immaginarsi una sorta di senescenza accelerata dei nostri antenati. In realtà
l'aumento della speranza di vita alla nascita è dovuto soprattutto al calo
della mortalità infantile: fino al XVIIIº secolo la probabilità di morire
entro i 10-15 anni era intorno al 50%[33], mentre la longevità non ha invece
subito variazioni così notevoli[34]. Dal punto
di vista del metodo c'è ancora da segnalare che le tecniche sviluppate dai
demografi per l'indagine retrospettiva delle popolazioni — in particolare
la ricostruzione delle famiglie — sono preziose per il genetista[35],
ad esempio nello studio delle caratteristiche recessive.
Se adottiamo un'ottica di tipo
sistemico[36] possiamo cercare le condizioni in base
alle quali un sistema demografico può configurarsi come omeostatico[37]. Il punto
di partenza è la mortalità, che possiamo considerare come esogena; né la convenienza
individuale né le norme culturali tendono infatti a rifiutare un abbassamento
di mortalità: sono allora nuzialità e fecondità — controllate in una certa
misura dal sistema di norme sociali — a dovere, per così dire, giocare di
rimessa. Come a fortiori ci aspettiamo
avvenga anche per gli altri animali, per la nostra specie le forze di eliminazione
sono probabilmente sempre state assai robuste. In più, la mortalità umana
è stata anche caratterizzata da picchi improvvisi e violenti. Questo può valere
per tutte le specie viventi (si pensi alle catastrofi naturali — pare ne sappiano
qualcosa i poveri dinosauri); ma proprio la superiore organizzazione sociale
e produttiva ha reso la nostra specie anche più vulnerabile a tali poco gradevoli
oscillazioni: se gli eventi bellici hanno avuto nel passato un impatto diretto
spesso modesto, è innegabile che l'affollamento in centri abitati e i contatti
dovuti ai commerci abbiano di molto favorito la comparsa di epidemie. L'Homo
sapiens è riuscito a surclassare le specie di predatori di grosse dimensioni,
che sicuramente più hanno turbato i sonni dei nostri antenati (i quali a loro
volta potevano allietarne i pasti); ma ha scoperto — come gli extraterrestri
ne La guerra dei mondi di Wells[38] — i nemici più grandi tra gli esseri viventi
più piccoli[39]
(oltre che in se medesimo[40]).
L'andamento della mortalità, per
tornare ai nostri sistemi omeostatici, ha dunque richiesto alla fecondità
non solo un livello sufficientemente alto per controbilanciare le perdite,
ma anche una grande flessibilità per potersi sincronizzare con andamenti estremamente
irregolari[41]. Esistono probabilmente delle dinamiche
strettamente biologiche che agiscono in questo senso, oltre a quelle che in
circostanze ordinarie temperano la fecondità rispetto al livello teoricamente
massimo. Studi sugli animali ipotizzano ad esempio una denatalità da sovrappopolazione;
spesso aumenta anche l'aggressività intraspecifica arrivando al cannibalismo,
oltre che a pratiche sessuali non finalizzate alla riproduzione, per arrivare
addirittura al suicidio collettivo[42].
L'umanità non sfugge necessariamente
a queste dure regole, sebbene possa mettere in atto strategie essenzialmente
culturali. Wrigley (pp.42-4) fornisce una panoramica delle soluzioni escogitate
da diversi gruppi umani oggetto di indagine antropologica: si va dalle interminabili
astinenze dei Djuka (per aspera ad astra) agli stratagemmi anticoncezionali degli Achehnese,
dalle deviazioni sessuali nei Tikopia agli infanticidi degli eschimesi Netsilik,
dei siberiani Ghiliachi e degli australiani Arunta (la lista continua con
Nootka, Thlinkeet, Rendille, Bangerang, Narrinyeri, Kurnai, Whatdhaving e
Motu Motu). Ma l'ammortizzatore principale, perlomeno nelle società prevalentemente
agricole dell'Europa post-medievale, era rappresentato dalle fluttuazioni
dell'età al matrimonio influenzata in particolare dai meccanismi giuridico-economici
relativi alla successione.
1.6 Fine della selezione naturale?
Sembra proprio che non tutto il
bene venga per giovare. Tra i frutti avvelenati che il calo della mortalità
relativo alla transizione demografica[43] ha recato
all'umanità, non ci sarebbe da annoverare solo la sovrappopolazione. Molti
paventano infatti che, dal punto di vista più propriamente evoluzionistico,
questo sviluppo possa averci allontanato dalla selezione naturale. Come risultato
dei progressi di medicina ed igiene, la riproduzione differenziale non si
baserebbe più, in particolare nei paesi a sviluppo avanzato, su differenze
di resistenza alla mortalità. Molto più peso avrebbe casomai la propensione
alla procreazione, che possiamo presumere influenzata maggiormente da variabili
socioeconomiche che non genetiche.
Steve Jones ha rilanciato una
polemica (cfr. l'articolo de laRepubblica del 13 dicembre
1999[44]) che non era certo nuova. Leridon
ad esempio (pp.92-4) metteva in rilievo come per queste ragioni[45] "the weight of biological factors,
therefore, has been considerably reduced". Perentorio Monod: "Il est évident qu'au sein de sociétés modernes, la dissociation est totale.
La selection y a été supprimée. Du moins n'a-t-elle plus rien de "naturel"
au sens darwinien du terme". Questo perché "l'intelligence, l'ambition, le courage, l'imagination sont certes toujours
de facteurs de succés dans les sociétés modernes. Mais de succès personnel, et non génétique[46], le seul qui compte pour l'evolution", mentre "il y a plus: à une époque encore récente, même dans les sociétés relativement
"avancées, l'elimination des moins aptes, physiquement et aussi intellectuellement,
était automatique et cruelle. La plupart n'atteignaient pas l'âge de la puberté.
Ajourd'hui, beaucoup de ces infirmes génétiques survivent assez longtemps
pour se reproduire"(p.206). In effetti in alcuni passaggi l'argomentazione, che rispecchia
i timori della demografia qualitativa britannica a cavallo tra ottocento e
novecento, non pare lucida come il resto dell'opera. Non fosse altro perché,
se veramente oggi questi danni genetici consentono comunque una vita quasi
normale, dobbiamo prendere atto del fatto che non costituiscono più una tara
grave[47]. L'attività antropica si configura né
più né meno come un cambiamento ambientale di altra origine — "naturale" verrebbe
da dire.
Inoltre mi chiedo quante lacrime
sia giusto versare sulla (presunta) fine della selezione naturale. Anzitutto,
secondo quale criterio giudicare la questione? Personalmente propendo per
la massimizzazione dell'utilità universale — non condivido perciò il reverenziale
rispetto per la natura (se ne parlerà oltre), né il passaggio logico dalla
scoperta dell'importanza della selezione darwiniana alla necessità di favorirne
l'operare[48]. In quest'ottica utilitaristica va
allora detto che la selezione naturale è un meccanismo con alcuni pregi (per
esempio una certa solidità) ma anche difetti evidenti: non solo essa è estremamente
rigida e lenta, ma soprattutto lastricata di sangue e sofferenze (cfr. Dawkins
in Appendice). Noi godiamo oggi del sacrificio di innumerevoli generazioni
di nostri (mancati) antenati, per quanto l'uniformità ambientale con il passato
ce lo permette[49]. Ma non possiamo
certo auspicarci di diventare noi stessi oggetto di tale inefficiente adattamento.
Inoltre non è neanche detto che
la pressione selettiva sia veramente diminuita. Ruse ad esempio lo nega (p.285)
"La conclusione perciò è che gli uomini
hanno alterato e continuano ad alterare il corso dell'evoluzione. Questo malgrado
molti si illudano [o paventino]
oggi che grazie alla moderna tecnologia l'evoluzione biologica dell'uomo si
è arrestata. La verità è che invece l'abbiamo accelerata[50]".
è poi chiaro che eventuali progressi dell'ingegneria genetica potranno rimettere
completamente in discussione i termini della questione[51].
Ciò che però rende per noi interessante
la controversia è che il demografo può dire più che una parola al proposito.
Procediamo con ordine:
ĀĀ dal punto di vista della mortalità, bisogna
considerare le componenti per causa e soprattutto per età. La morte dopo l'età
feconda non ha conseguenze (dirette, perlomeno[52])
sull'ereditarietà (cfr. Cliquet p.202). Da qui forse le sofferenze fisiche
della condizione anziana, sottoposta solo assai marginalmente a selezione
naturale[53]. Vediamo allora che in effetti le cause
di morte infantili e giovanili si sono assai ridotte, anche se (specie le
seconde) non certo annullate: alcolismo e suicidio giovanili, AIDS, droga,
incidenti stradali sono esempi abbastanza evidenti. Di fronte a molte nuove
insidie in effetti manca una copertura genetica[54], quindi è immaginabile un graduale orientamento
in questo senso. L'ottimismo relativo al calo della mortalità può rivelarsi
d'altronde mal riposto alla luce dell'aumento degli agenti inquinanti e mutageni.
Ci siamo mai chiesti se l'aumento della speranza di vita reggerebbe ancora,
se potessimo retrodatarne il conto dalla nascita al concepimento? Da quanto
si sospetta sulla diminuzione della fertilità umana[55] c'è da pensare
che si potrebbe avere qualche sorpresa. Se così fosse la selezione non avrebbe
addolcito (nella misura che si crede, almeno) la propria morsa; anche se c'è
da convenire che l'ipotetico anticipo della mortalità infantile all'età prenatale
sia comunque vantaggiosa (nell'ottica piacere/dolore) per i nostri conspecifici[56].
ĀĀ per quanto riguarda la fecondità, va
detto anzitutto che i demografi potrebbero aiutare a verificare fino a che
punto è vero che oggi i meno adatti si riproducano maggiormente, oltre che
sopravvivere: per molti difetti genetici questo mi pare poco verosimile. è
invece certo[57] che l'aumentata mobilità abbia diminuito
il grado di inbreeding, col simpatico risultato di ridurre anche l'incidenza fenotipica
delle tare recessive (vedi Lucchetti e Soliani p.163, Cliquet pp.187-8 e 205).
Un eventuale spostamento della fecondità verso le età meno giovani avrebbe
poi l'effetto di aumentare il pericolo di aberrazioni cromosomiche, ma sul
lunghissimo periodo anche quello — suppongo caro ai selezionisti radicali
— di estendere la copertura genetica ad età più avanzate[58].
ĀĀ una delle conseguenze più eclatanti della
transizione demografica, più precisamente delle differenze di tempi e modi
nel compierla[59], è quella
di aver reso prevalente la selezione demica rispetto a quella individuale[60]. Da questo punto di vista si può ammettere
che i fattori genetici di selezione diventino marginali rispetto alle condizioni
socioeconomiche tra le diverse aree del mondo. Anche qui, è possibile che
una certa importanza assumano in futuro le tecniche artificiali di riproduzione
(cfr. n.63 più sotto) — che potrebbero ridurre il divario
di fecondità tra le popolazioni ricche e quelle povere.
Ecco allora che i demografi possono
raccogliere e analizzare i dati, avanzare previsioni, studiare le interconnessioni
tra i fenomeni. Possono insomma dare sostanza e misura alle diverse ipotesi
anche su questo terreno.
[1]
Ringrazio Carlo Maccheroni per i preziosi consigli.
[2]
Su questo e altri argomenti toccati in quest'introduzione si veda Federici
1987.
[3]
Cfr. Keyfitz, ripreso da Scardovi (1985): "Tra
l'analisi biologica e quella sociale della popolazione si apre un'ampia
area di ignoranza. Come tanti altri scienziati sociali, i demografi si sono
preoccupati di evitare la biologia perché ritenuta non rilevante ai loro
fini, o addirittura pericolosa.
(...) I manuali di biologia incorporano brevi trattazioni di demografia.
Ma questa attenzione non è reciproca." Continua poi Scardovi: "è vero. Ed è vero anche che se si vuole infastidire un demografo — salvo
alcune belle, luminose eccezioni — si deve parlargli di biologia delle popolazioni,
di genetica delle popolazioni: vale a dire di scienza". Anche Cliquet (p.183) si riferisce
a Keyfitz: "demography appears to
be of much greater importance [per la genetica di popolazione] than is often thought, even in the demographic
community itself. Whereas demographic variables are routinely included in
bio-anthropological and population genetic work (e.g. Adams et al., 1990;
Cavalli-Sforza & Bodmer, 1971), demography only occasionally deals with
these interrelationships (e.g. Keyfitz, 1984)". Rincara la dose,
allargando anche il campo, Van der Berghe (p.98): "... gli ostacoli intellettuali che gli studiosi
di scienze sociali erigono fra se stessi e la loro comprensione del pensiero
biologico evoluzionistico. L'onere del riavvicinamento ricade chiaramente
su di essi. Sono essi a dover tornare alla fraternità scientifica. Sono
essi che dovranno gettar via i paraocchi intellettuali che si erano autoimposti."
[4]
Possiamo trovare un po' esagerato l'entusiasmo di Pietro Verri ("gli uomini forse più benemeriti del genere
umano sono quelli che si occupano di storia naturale" — nella mostra
Pietro Verri e la Milano dei Lumi, Milano
gennaio-marzo 1998, senza indicazione della fonte bibliografica). Più facile però concordare con quanto è stato
autorevolmente sostenuto, "la biologie
est-elle, pour l'homme, la plus signifiante de toutes les sciences; celle
qui a déjà contribué, plus que toute autre sans doute, à la formation de
la pensée moderne, profondément bouleversée et définitivement marquée dans
tous les domaines: philosophique, religieux et politique, par l'avènement
de la théorie de l'Evolution" (Monod, p.11).
Disconoscere la rilevanza della biologia sarebbe
a mio modo di vedere una grave miopia intellettuale, anche se comprensibile
alla luce degli abusi che spregiudicate dottrine sociologiche e soprattutto
criminali dittature ne hanno fatto nel passato.
[5]
Questo non vale per tutti, naturalmente (le belle,
luminose eccezioni cui si riferisce Scardovi 1985). Né vale necessariamente
per il passato, quando un approccio di tipo naturalistico era assai più
diffuso. Si veda ad esempio I fattori
biologici dell'ordinamento sociale di Livio Livi (1936), poi ristampato
come primo volume del Trattato di
Demografia, che contiene tra l'altro una rassegna del pensiero pre-wilsoniano
(chiamiamolo così) da Darwin in poi. Nella prefazione è scritto: "Le forme associative possono considerarsi più o meno come conseguenza
di taluni caratteri di specie. Sicché (...) mi confermai nell'opinione che
non si poteva, per lo studio delle società stesse, tagliare ogni collegamento
col mondo naturale. Mi convinsi anzi che i fatti del mondo animale potevano
dar molta luce allo studio della società umana, e che considerando l'uomo
dal punto di vista zoologico si poteva avere una buona guida per la comprensione
dell'essenza della società e delle forze che la tengono unita" (pp.LXIX-LXX).
Su questo vedi Scardovi 1994 (specialmente pp.191-9).
[6]
Scardovi (1985) parla della demografia come di una "virtuale koinè euristica"; Federici, tra gli altri, la definisce "disciplina-ponte tra le scienze naturali e
le scienze sociali"(p.20). Diversi contributi su questo punto sono menzionati
in de Bruijn, pp.1-3.
[7]
"Il più certo modo di celare agli
altri i confini del proprio sapere, è di non trapassarli" (Leopardi,
Pensieri, LXXXVI).
[8]
Lucchetti e Soliani accennano, a proposito dell'analisi dei cognomi, alla
possibilità di superare "la distinzione,
troppo a lungo mantenuta, tra l'approccio biologico, demografico e socio
culturale nello studio delle popolazioni umane e della loro storia ed evoluzione"
(p.175). Anche il contributo di Scardovi del 1994 perora la causa di una
maggiore apertura interdisciplinare della demografia, in particolare verso
la biologia — un punto di vista condivido ampiamente. Rimando dunque al
suo intervento per una trattazione più ampia di questo tema.
[9]
Non insisterò invece, se non di passaggio, sul piuttosto scontato presupposto
biologico della demografia.
[10]
Il termine fertilità indica il
potenziale fisiologico di procreazione, mentre fecondità si riferisce all'effettivo numero di figli. Si noti però
che in inglese il significato di fecundity
e fertility è invertito rispetto
alle lingue latine, con immediato effetto su molte traduzioni.
[11] "Probably for historical reasons, demography developed using almost exclusively
concepts and tools developed by statisticians. Population genetics on the
other hand, first went through a period of predominant use of statistical
analysis (notably under the influence of R.Fisher); then it grew mainly
by using probabilistic thinking (Sewall Wright and, most of all, M.Kimura).
It would no doubt be useful for there to be a similar evolution of demography
towards a generalized use of the probabilistic language" scrive Jacquard
(p.31), che poi aggiunge "Demographers
moreover pay attention to the mean of a studied parameter, while geneticists
note rather its variance. This discrepancy is especially clear about fertility
[cfr. n.45]". Su quest'ultimo aspetto vedi per es. Gallino p.XXIII.
[12]
Continua poi (pp.206-7): "L'intersezione
tra genetica di popolazioni e demografia scientifica attiene dunque anche
ai paradigmi teorici: e sono paradigmi statistico-probabilistici; come la
genetica di popolazioni indaga il modificarsi delle frequenze statistiche
dei geni, così la demografia osserva il rinnovarsi e il distribuirsi dei
portatori di quei geni: e sono distribuzioni statistiche".
[13]
La più intuitiva delle misure sintetiche della fecondità è il TFT
(tasso di fecondità totale), che si costruisce semplicemente sommando i
tassi di fecondità specifici per età, e che dà come risultato il numero
di figli di ambo i sessi che una coorte di donne avrebbe in assenza di mortalità.
Per il tasso netto di riproduzione, assai importante in demografia, il calcolo
è invece effettuato separatamente per sesso (di regola quello femminile),
e tiene conto anche dell'effetto della mortalità: il numero delle madri
potenziali ad ogni età viene infatti decurtato dei decessi; l'ammontare
finale della discendenza femminile è pertanto minore che non nell'ipotesi
di assenza di mortalità (la misura sarebbe in quest'ultimo caso il tasso
lordo di riproduzione R). La formula è: ĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀ(1)
Le quantità fx sono i tassi di fecondità
specifici (numero di figli avuti in media nei dodici mesi di età x compiuta), mentre a e b-1 sono
rispettivamente l'età minima e massima del periodo fecondo (convenzionalmente
stabilite in 15 e 44 o 49 anni); k è infine il rapporto tra i sessi alla nascita (circa 105 maschi
per 100 femmine). Tratti dalle tavole di mortalità (che hanno riferimento
ad una generazione, anche fittizia) sono poi i sopravviventi all'età esatta
(lx) e gli anni vissuti nell'intervallo
(Lx, pari convenzionalmente
alla media aritmetica tra lx
ed lx+1). Il rapporto
Lx/l0, equivale alla
probabilità di sopravvivenza dalla nascita fino all'età compiuta x (equivalente in media all'età esatta
x+0,5); è questa probabilità a ridurre
la fecondità effettiva nel corso degli anni in ragione della mortalità.
Se la mortalità fino all'età b
fosse nulla, gli Lx
sarebbero tutti uguali ad l0:
la probabilità, uguale ad uno, sparirebbe dalla formula e quindi R0 coinciderebbe con R (se non tenessimo conto neanche del k, la formula sarebbe quella del TFT).
[14]
Cfr. Bodmer e Cavalli-Sforza, II, pp.58-61, par. La cinetica della selezione darwiniana, e Lucchetti e Soliani, §3.2.
[15]
Si intende qui quello che Wilson chiama "altruismo
a oltranza in contrapposizione con l'altruismo moderato: il primo è basato
su particolarissimi tipi di catena genetica tramite l'aiuto ai parenti e
il secondo consiste nella reciprocità — il ben noto contratto sociale che
governa tanta parte del comportamento umano" (Wilson-Harris, p.153).
Per una definizione si veda Gallino p.XV (che cita Barash): "Nel linguaggio della sociobiologia il termine altruismo non ha alcuna
connotazione morale, né presuppone un elevato livello di coscienza: è semplicemente
«un atto che riduce il successo riproduttivo personale di chi lo esegue
mentre aumenta il successo riproduttivo di altri»".
[16]
Cfr. Melotti p.78, che cita Maynard Smith. Cfr. anche Gallino p.XI.
[17]
Questa procedura è per esempio adombrata in Van der Berghe (pp.109-10):
"Si presume, per esempio, che i genitori i quali
hanno superato l'età della riproduzione siano più altruisti verso i figli
in grado di riprodursi di quanto non lo siano questi verso di loro, quantunque
il loro coefficiente di parentela sia identico".
[18] L'operazione è in verità ridondante: se nelle
formule presentate in questo paragrafo lasciassimo a tutto l'arco di vita
l'estensione delle sommatorie relative alla fecondità (scrivendo w-1 in luogo di b-1), il risultato non cambierebbe: daĀĀ bĀ in poi, infatti, tutti gli fx si considerano nulli.
[19]
Per una trattazione di tutti questi punti (altruismo, idoneità complessiva,
coefficiente di parentela biologica) vedi Melotti 1986, pp.76-8.
[20]
Vi sarebbero delle puntualizzazioni da fare dal punto di vista demografico.
Nella formula considero gli Lx
(discorso analogo può farsi per gli fx)
come relativi ad un individui, mentre essi hanno normalmente riferimento
collettivo. L'ipotesi implicita è allora quella di applicare all'individuo
in questione una tavola di mortalità, anche ideale: ciò che in realtà interessa
non è la procedura di costruzione della tavola di mortalità, bensì disporre
almeno in linea di principio di un modo per quantificare il tempo da vivere
— che può essere stimato anche soggettivamente.
[21]
Riassumendo, la formula-base dell'altruismo si limita perciò a calcolare
se Āè maggiore o minore di uno. Dinamizzare
rispetto all'età significherebbe aggiungere le speranze di sopravvivenza,
arrivando a: ĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀ(3)
(gli Lx
dovrebbero venire divisi per la radice, che vale 10n, della tavola
di mortalità ¾
cfr. nota 13¾ ma si semplifica tra numeratore e denominatore).
Possiamo poi estendere ulteriormente la (2), ad esempio
tenendo conto delle probabilità stimate a priori di salvare i consanguinei
e di perdere laĀ vita in conseguenza
del tentativo, il cui esito adesso non consideriamo più certo. Se p è la probabilità di sopravvivere (escludiamo
l'eventualità di ferite o mutilazioni) al singolo evento analizzato, avremo
allora: ĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀ(4)
L'ipotesi è che, in assenza del nostro intervento
le pi sarebbero state tutte nulle
e pego uguale ad uno.
[23]
In realtà tutte le popolazioni si allontanano dall'ideale un po' rousseauiano
di "fecondità naturale" (cfr. de Bruijn, pp.8-9), che dovrebbe rispecchiare
la pura fertilità.
[24]
Per possibili effetti distorsivi dal punto di vista selettivo si veda il
par. 1.6.
[25]
Un discorso parallelo era già stato fatto da Lorenz ne L'aggressività (vedi il cap.XIII): all'evoluzione degli strumenti
di offesa non avrebbe corrisposto uno sviluppo nelle inibizioni istintive
all'aggressione intraspecifica. In questo senso anche la ricerca di Dave
Grossman, On Killing, recensita
da Cristina Mochi.
[26]
L'istituzione del "matrimonio riparatore" tende per esempio a fare entrare
stabilmente in fase riproduttiva i più fertili (o forse i meno avveduti).
Questo vale in particolare nelle società tradizionali, nelle quali più stretto
è il legame precocità del matrimonio/discendenza finale.
[28]
A dire il vero questo sarebbe possibile già da tempo. Vi allude con nonchalance Harsanyi 1994: "Prendiamo ad esempio il sesso come variabile«immutabile»
(trascurando gli esempi di cambiamento di sesso)..." (p.152).
[29]
Una rassegna di contributi demografici allo studio della mortalità è in
Soliani e Lucchetti, §5.
[30]
Sarebbero possibili innumerevoli applicazioni, per esempio con attenzione
a variabili di carattere ambientale, quali l'esposizione a fattori inquinanti
o la densità di popolazione.
[31]
Per almeno i primi tre concetti è possibile dare definizioni formali. Relativamente
all'anno t avremo:
Età media al decesso: ĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀ(6)
In riferimento ad una determinata tavola di mortalità
abbiamo invece la speranza di vita alla nascita:
ĀĀĀĀĀĀĀĀĀĀ
(7)ĀĀĀĀĀĀĀĀĀĀĀĀĀ ĀĀĀĀĀĀĀĀĀĀĀ; alla generica età x vale: ĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀĀ(8).
I simboli rappresentano l'ammontare delle classi
di età (Px) e dei i
relativi decessi (Mx)
nella popolazione reale; a questa si contrappone la popolazione stazionaria
associata alle tavole di mortalità (per lx
e Lx si veda la nota
13).
Per la longevità può farsi invece riferimento al
cosiddetto punto di Lexis, ovvero
alla moda (questo termine dal sapore frivolo indica in statistica la classe,
qui di età, con massima frequenza) dei decessi in età adulta; Lexis pensava
di poter leggere nel grafico dei decessi per età — fatta astrazione da quelli
infantili e accidentali — una sorta di curva gaussiana con scostamenti casuali
dall'età normale alla morte. Chi preferisca riferimenti più umanistici potrà
ricorrere al Dialogo di un fisico
e di un metafisico: "[Metafisico] ...
Io so che oggi i vostri pari tengono per sentenza certa, che la vita, in
qualunque paese abitato, e sotto qualunque cielo, dura naturalmente, eccetto
piccole differenze, una medesima quantità di tempo, considerando ciascun
popolo in grosso." (Leopardi, Operette
morali). Sulle ipotesi biologiche concernenti l'invecchiamento,Ā cfr. Soliani e Lucchetti §3.
[32]
Molto inferiore, dunque, a quella dei paesi più poveri di oggi.
[33] Già osservava Hume: "Nature has rendered human infancy peculiarly
frail and mortal; (...) The half of mankind die before they are rational
creatures" (Of the Immortality
of the Soul, p.95, nei Posthumous
Essays).
[34]
Per quanto anch'essa si sia allungata. Dal punto di vista sociologico è
interessante la tesi di Ariès (p.65): "Se
durante il vecchio regime si entrava nella vita più giovani, se ne usciva
anche meno vecchi: e ciò a causa della morte, senza dubbio più frequente,
ma soprattutto perché ci si ritirava. Arrivato sulla cinquantina, l'attività
dell'uomo rallentava. Egli faceva già la figura del vecchio che i giovani
ingannavano e sberleffavano". Ariès accenna anche ai "vecchioni di Molière". Dovendo portare prove letterarie, aggiungerei
l'illuminante IIº sonetto di Shakespeare:
When forty winters shall besiege thy
brow,
And dig deep trenches in thy beauty's
field,
Thy youth's proud livery so gaz'd on
now,
Will be a tatter'd weed of small worth
held:
Then being ask'd, where all thy beauty
lies,
Where all the treasure of thy lusty
days;
To say within thine own deep sunken
eyes,
Were an all-eating shame, and thriftless
praise.
How much more praise deserved thy beauty's
use,
If thou could'st answer this fair child
of mine
Shall sum my count, and make my old
excuse
Proving his beauty by succession thine.
This were to be new made when thou
art old,
And see thy blood warm when thou feel'st
it cold.
Rivolgetevi oggi ad un quarantenne in questi termini
(parlo anche a titolo personale), e avrete rotto un'amicizia.
[35]
Cfr. Jacquard, p.30 e Bodmer e Cavalli-Sforza v.I, pp.39-66. Segnalo anche
l'articolo Tra scienza e genealogia,
che riporta una ricerca sul morbo di Alzheimer che è stata spinta sino alla
prima metà del XVIIº secolo.
[36]
Questa impostazione, più affine all'indagine biologica, non è certo l'unica
possibile: possiamo ad esempio immaginare un approccio di tipo economico
— l'ottica sarà allora quella della convenienza del singolo; oppure uno
di tipo sociologico, incentrato sulla dinamica culturale, in particolare
dei valori, e sullo studio della cogenza di questi ultimi sui comportamenti
riproduttivi (se non anche di sopravvivenza). Hobcraft
e Kiernan, p.31, riportano l'affermazione di Duesenberry "economics is about choice; sociology is about lack of choice".
[37]
Nelle popolazioni storiche possiamo assumere la stazionarietà demografica
come condizione di equilibrio del sistema, anche se ciò rappresenta una
semplificazione: anzitutto va osservato che il trend epocale della popolazione
umana è stato positivo, seppure quasi sempre in misura impercettibile e
con parentesi anche durature di regresso; Livi Bacci 1981, p.114, mette
inoltre in rilievo come stabilità strutturale non significhi necessariamente
stabilità demografica. Un approccio evoluzionistico può infine ispirarci
un certo scetticismo verso un rigido utilizzo del concetto di equilibrio
(si veda Boulding, p.72).
[38]
Possiamo pensare anche all'epico duello di stregoneria de La spada nella roccia.
[40] Dovendo aggiungere l'ipse dixit all'evidenza, possiamo ricorrere a Monod (p.204): "Dominant désormais son environnement, l'Homme
n'avait plus devant soi d'adversaire sérieux que lui-même. La lutte intraspécifique
directe, la lutte à mort, devenait dés lors l'un des principaux facteurs
de sélection dans l'espèce humaine. Phénomène extrêmement rare dans l'évolution
des animaux."
[41]
Questo tipo di interpretazione causale è l'inverso di quella malthusiana
tradizionale, secondo la quale le forze di eliminazione si incaricano di
riportare l'ammontare della popolazione, dilatato da una natalità eccessiva,
a livelli sostenibili dalle sussistenze: qui è infatti la mortalità a venire
considerata esogena, mentre la fecondità deve adeguarvisi.
[42]
Reynolds scrive (p.160, in Furcht 1985, p.120): "In certe specie di roditori, particolarmente topi campagnoli, alcuni cambiamenti
fisiologici del sistema riproduttivo (ad esempio, il riassorbimento del
feto) si associano a condizioni di sovraffollamento. Sembra che queste condizioni
in molte specie di roditori, come pure in molti altri mammiferi, si accompagnino
a situazioni di notevole attenzione emotiva, con l'aggressività come fatto
concomitante, comune nei ratti e nei topi, e l'uccisione dei piccoli, che
vengono poi divorati, nei ratti." E poi prosegue (p.167, in Furcht 1985,
p.121): "Quanti più animali vengono
raggruppati in un determinato spazio, cioè quanto maggiore è la densità
di popolazione, tanto più elevato sarà il livello globale [pro-capite,
intende] di secrezione adrenalinica
e noradrenalinica da parte di tutti gli animali, specialmente di quelli
subordinati. Da un punto di vista comportamentale, si vede che la crescente
densità si correla con una crescente aggressività da parte degli animali
dominanti, mentre i subordinati tendono a scappare o si rannicchiano per
la paura. A questo stadio, la riproduzione può cessare, i giovani possono
venire divorati e, in generale, l'intera serie di cambiamenti può essere
vista come meccanismo per fare selettivamente spazi.". Vedi anche Lorenz,
p.300.
[43]
Con "transizione demografica" si intende il passaggio ad un regime demografico,
proprio della nostra epoca, caratterizzato da bassi livelli di natalità
e mortalità.
[44]
Vi si legge: "Il caposaldo numero
uno del darwinismo — e cioè l'esistenza di un brutale processo di selezione
naturale in base al quale sopravvivono solo gli esseri più forti — «non
è più presente nella società moderna. In altri tempi il 'debole' sarebbe
morto prima di avere figli. Adesso invece si riproduce con altrettanta efficienza».
La medicina gli consente non solo di vivere a lungo ma di colmare quasi
ogni handicap. La vita urbana con il suo vorticoso gioco delle coppie ha
portato ad un generale livellamento 'medio' nella qualità dei geni".
Per una più sfumata conferma della parte di questa tesi relativa alla mortalità,
si veda Cliquet pp.201-5. Una valutazione degli effetti dell'odierno meccanismo
di formazione delle coppie sempre in Cliquet, §3.1.
[45]
Si riferisce anche, dal lato della fecondità, alla riduzione nella varianza
del numero dei figli tra le coppie. Sulla fecondità (e sugli effetti dell'aborto
terapeutico sul pool genico) si veda Cliquet §3.2.
[46]
Possiamo in verità aspettarci qualche vantaggio dal lato della sopravvivenza
(misurabile ad esempio in termini di riduzione delle probabilità di morte
per età qx), benché l'ipotizzabile concentrazione
nelle età anziane lo renda poco significativo dal punto di vista della selezione.
L'ipotesi di Monod sulla fecondità, ragionevole, è che il successo sociale
negli animali (quelli che vivono in gruppo, ovviamente) si traduca di norma
in successo sessuale e perciò in una più numerosa discendenza. Per gli esseri
umani il successo sociale (che comprende anche quello economico), anche
comportasse un maggior successo sessuale, non implicherebbe però un maggior
numero di figli (cfr. sez.1.2). Si veda però ancora il §3.2 di Cliquet, in particolare
il riferimento all'ipotesi eugenica (p.200) "In some countries the the earlier negative association between fertilità
and SES-indicators show the first signs of a possibile reversal, confirming
Osborn's (1952; 1968) eugenic hypothesis, stating that in a situation of
free choice the distribution ofĀ births
will favour more children being born in the most favourable home environments.
This implies that the end of the demographc transition might be characterised
by a positive association between reproductive fitness and socially valuable
traits" e, nel seguito, alle relative conferme empiriche.
[47]
Di questo parere anche H.Harris, cfr. pp.7-8.
[48]
Newton non si è certo buttato dalla torre di Londra dopo avere scoperto
la gravitazione universale. Su questo mi pare chiaro Ruse, che esprime anche
qui un parere equilibrato (pp.271-7); in particolare, dopo aver menzionato
l'errore naturalistico rilevato da Hume nel
Treatise, evidenziato proprio
a proposito dell'etica evoluzionistica da G.E.Moore nel 1903 (vi si soffermano
molti testi, cfr. ad es. Musacchio p.20 e Rollin p.24; ma vedi soprattutto
Boccara 169-70 e, ancora più diffusamente, Ricken pp.49-56), osserva: "Ora, è abbastanza chiaro che gli etici evoluzionisti
commettono l'errore naturalistico. Infatti passano da «Così è il mondo»
(a causa dell'evoluzione) a «Così dovrebbe essere il mondo» (e perciò bisogna
aiutare l'opera dell'evoluzione)" (p.273); cfr. ancheScarpelli p.6 e Popper p.100. Nel merito
dell'etica evoluzionistica si veda infine Engelhardt pp.48, 196 e 275.
[49]
Ricorda Engelhardt, a proposito dell'identificazione delle malattie: "Noi siamo il prodotto di forze selettive cieche,
le quali, se hanno avuto successo, ci hanno adattato ad ambienti nei quali
è possibile che non viviamo più" (p.196) e aggiunge poi (p.197): "si deve notare ancora un'altra difficoltà nell'appellarsi
alle condizioni della natura o ai risultati dell'evoluzione. Un appello
del genere rende i propri giudizi (...) ostaggio del passato".
[50]
Si riferiva esplicitamente all'incremento di stress e tensione (che determinano
un rilevante mutamento nel valore adattativo del genoma), agli scambi di
agenti patogeni tra popolazioni diverse grazie alla mobilità, alle conseguenze
dell'urbanizzazione, all'aumento delle sostanze mutagene. Sulla possibilità
che la transizione demografica non abbia diminuito la pressione selettiva,
cfr. Cliquet pp.197-8.
[51] Devo incassare l'appellativo di demi-savant che Monod (p.207) appioppa
a chi diffonde quest'illusione, visto che "l'echelle
microscopique du génome interdit pour l'instant et sans doute à jamais de
telles manipulations". Anche stavolta Monod mi pare un pochino
troppo — come si dice in francese — tranchant.
Ruse infatti (pp.285-6) è più possibilista, e Rollin dedica il suo libro
a possibili sviluppi di questo genere di tecniche. è in fondo un lato bello
della scienza che, nonostante Verne e altri profetici autori di fantascienza,
sia impossibile prevederne gli sviluppi (si veda in questo senso Popper,
Prefazione a Miseria dello storicismo).
[52]
Possiamo immaginare qualche tenue legame da un lato con i benefici (sfruttamento
delle residue potenzialità produttive, trasmissione dell'esperienza e protezione
della prole — i nipoti, magari) e dall'altro con il peso che il mantenimento
dell'anziano (inteso qui come individuo al di là dell'età fertile) comporta
per le generazioni più giovani.
[53]
Un'ipotesi più sofisticata in Nesse e Williams: "Nel 1957 uno di noi (Williams) ipotizzò che i geni che causano l'invecchiamento
e la morte potessero comunque essere favoriti dalla selezione naturale qualora
avessero effetti vantaggiosi per i più giovani, quelli cioè che sono maggiormente
sottoposti a pressione selettiva.[Seguono esempi relativi al metabolismo
del calcio, alla gotta, al sistema immunitario: per "i più giovani" si intende
il medesimo individuo in età giovane]" Gli autori aggiungono poi però:
"è anche possibile, naturalmente, che la maggior parte dei geni che causano
l'invecchiamento non porti benefici in alcuna fase della vita; in natura,
questi geni potrebbero semplicemente non avere sulla capacità riproduttiva
effetti tali da essere eliminati per selezione naturale." Si vedano
anche Cliquet p.190 ed Engelhardt p.197.
[54]
Facciamoci ancora aiutare da Ruse (p.254): "Quando
si tratta di fobie l'uomo mostra la sua storia evoluzionistica. Per esempio,
molti di noi rabbrividiscono di fronte ai serpenti, fenomeno di ovvio significato
adattativo. D'altro canto, malgrado i frequenti incidenti, non si ha la
stessa reazione di fronte ai coltelli, alle prese di corrente e ad altri
oggetti pericolosi che si trovano in casa". Si tratta poi di allargare
il discorso a coperture anche non comportamentali verso ad esempio nuovi
agenti virali, inquinamento, fumo di sigaretta, alcool, droghe, alimentazione
eccessiva e insana.
[55]
Per esempio riduzione in numero e motilità degli spermatozoi. Mi pare ragionevole
considerare l'ipotesi che i mutamenti ambientali responsabili di tale supposta
diminuzione possano anche riflettersi sulla mortalità intrauterina (cfr.
n.82), che
a sua volta – se molto precoce – può essere difficilmente distinguibile
da una minor fertilità. Sulla valenza selettiva della mortalità intrauterina
cfr.Cliquet p.190.
[56]
Scrive Engelhardt a p.251: "è estremamente improbabile che i feti, con
i loro lobi frontali non completamente sviluppati né completamente connessi,
siano in grado di provare sofferenza, anche se sono in grado di provare
dolore. La sofferenza richiede il riconoscimento del dolore come qualcosa
di minaccioso, di negativo, da evitare.Ā
La sofferenza richiede qualcosa di più di una semplice proprietà
dolorosa. Richiede un riconoscimento di quella proprietà, di quella sensazione,
come qualcosa di sbagliato, di dannoso e di nocivo. è discutibile se i feti
all'inizio della gestazione provino anche solo dolore."
[57]
Cfr. Cliquet pp.194-6 per un parere più articolato.
[58]
Cfr. Bodmer e Cavalli-Sforza (p.56) per il caso della schizofrenia. Tra
i mali a forte componente ereditaria che insorgono tardivamente, merita
di essere ricordata almeno la corea di Huntington (cfr. H.Harris p.69, e
Milano pp.60 e 115).
[59]
La transizione comporta quasi sempre un periodo iniziale di forte incremento
della popolazione, perché inizia di norma con una discesa di mortalità,
cui la fecondità si adegua con un ritardo che può essere anche di diversi
decenni.
[60]
"La selezione darwiniana è talvolta
chiamata selezione intragruppo, poiché agisce sugli individui di un gruppo.
Può esistere anche una selezione naturale che favorisce un gruppo rispetto
a un altro, nel qual caso si parla di selezione intergruppo. Si tratta di
una selezione che non necessariamente si riferisce a una competizione,a
un conflitto diretto tra i gruppi. Può verificarsi semplicemente che certi
gruppi abbiano un incremento di popolazione maggiore di altri (selezione
demica)" (Bodmer e Cavalli-Sforza, II, pp.62-3).